
ECCEZIONI ALLE LEGGI DI MENDEL
L’eredità intermedia
È un’eredità senza dominanza. Sui testi di genetica è famosissimo l’esempio dell’incrocio tra fiori rossi e fiori bianchi. Nella prima generazione F1, si ottengono solo fiori rosa, ma l’incrocio F2 dà il 25% di fiori rossi, il 50% di fiori rosa e il 25% di fiori bianchi.
In questo caso abbiamo quindi tre fenotipi e tre genotipi.
AA = rosso
Aa = rosa
aa = bianco
AA x aa = 100% Aa
Aa x Aa = 25% AA, 50% Aa, 25% aa.
Forse, però, non ha più senso indicare i geni con lettere maiuscole e minuscole. Solitamente si differenziano con segni in pendice (es. Ar e Ab). Esempi di questo tipo d’eredità sono presenti nei polli, nei bovini e in diverse specie di uccelli. Nel cane non c’è nulla di scientificamente provato (un’ipotesi potrebbe essere l’orecchio a tulipano, intermedio tra l’orecchio eretto e quello semipendente). In ogni caso, a questo tipo di incrocio, possiamo assimilare moltissime mutazioni quantitative, purché non si voglia entrare troppo nel dettaglio.
Per esempio, dall’accoppiamento di un cane tigrato scuro (molto nero, rispetto al rosso) e uno tigrato chiaro (molto rosso rispetto al nero), posso aspettarmi di ottenere tigrati di gradazione intermedia. Reincrociandoli tra loro, posso aspettarmi di avere il 25% di chiari, il 50% di intermedi e il 25% di scuri, valutando la tigratura nel complesso, e facendo una suddivisione arbitraria. In realtà non c’è un confine netto che divide il chiaro dall’intermedio e l’intermedio dallo scuro.
La distribuzione 25 – 50 – 25 non è altro che una rudimentale campana di Gauss.
L’eredità quantitativa
Sempre mantenendo il riferimento al colore, per facilità di comprensione, pensiamo ancora alla tonalità del colore fulvo. Alcuni sono chiari e altri scuri, con tantissime scalature intermedie di tonalità. Lo stesso discorso vale per i roani che possono essere quasi bianchi con poche punteggiature o quasi brizzolati, con peli scuri che prevalgono su quelli bianchi.
Nella morfologia ci sono tantissimi esempi: la taglia, la lunghezza degli orecchi, la lunghezza della coda, il numero dei denti. La genetica attribuisce questo tipo di eredità all’interazione contemporanea di più geni (poligeni o multipli) ognuno dei quali porta un suo contributo maggiore o minore al manifestarsi di un certo fenotipo. Il risultato finale è dovuto all’azione cumulativa di tutti i geni. Quanto più numerosi sono i poligeni mutanti tanto più il fenotipo discosterà dall’esemplare non mutato. Un cane grigio con pochi poligeni schiarenti sarà pressoché simile a un nero. Se alla stessa mutazione abbiniamo un numero altissimo di poligeni, avremo un cane quasi bianco.
Un esempio di trasmissione poligenica è il colore più o meno scuro del golden retriever. Tutti i golden retriever hanno lo stesso corredo genetico per quanto riguarda il colore: sono tutti fulvi recessivi. Tuttavia, in alcuni si manifesta un colore rosso carico (comunque mai mogano), in altri un colore molto sbiadito (comunque mai bianco). La tonalità dipende da una serie di geni modificatori. Per semplicità ipotizziamo si tratti di sei poligeni (potrebbero essere molti di più)
Dall’accoppiamento di due cani con le due espressioni estreme del mantello (quindi uno quasi crema e uno fulvo carico) avremo la totalità della prole con colore intermedio. Infatti il cane dal mantello più scuro possiede sei geni modificatori del tipo scuro e ne trasmette tre alla prole. Il partner dal mantello chiaro possiede sei geni modificatori di tipo chiaro e ne trasmette tre a sua volta. I cuccioli avranno tre geni modificatori scuri e tre chiari.
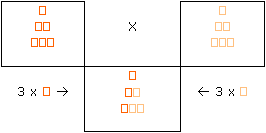
Nel reincrocio otteniamo numerose tonalità. La tonalità intermedia è la più frequente, mentre le tonalità estreme (in entrambe le direzioni) sono più rare.
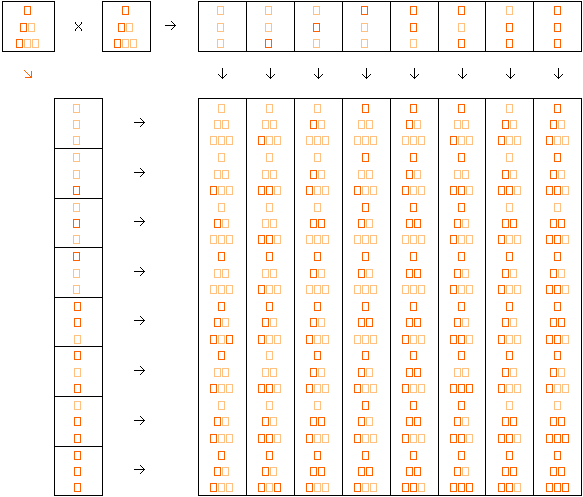
L’esempio proposto è molto semplificato. In realtà, in questo tipo di eredità, i geni che interessano un carattere sono solitamente molti di più. Il risultato della loro interazione può essere valutato solo statisticamente e su numerose cucciolate, attraverso una curva di variabilità Gaussiana).
I metodi di studio dei caratteri quantitativi però, tutto sommato, sono semplici, perché si basano sul fatto che i singoli poligeni seguono un modello mendeliano di trasmissione ereditaria.
La penetranza incompleta
Anche in questo tipo di eredità non vi è prevalenza di un carattere sull’altro. A differenza dell’eredità intermedia, in questo caso, vi può però essere una prevalenza di uno sull’altro. Per esempio la pezzatura (macchie bianche) è ritenuta, da molti, soggetta a questo tipo di trasmissione. Il mantello solido prevale su quello pezzato e si comporta quasi come dominante. I portatori di pezzatura sarebbero però spesso riconoscibili perché spesso presentano piccole macchie bianche al petto, alle zampe o al muso.
Ma la caratteristica più interessante di questo tipo di eredità è comunque la diversa espressione della mutazione tra soggetti con lo stesso patrimonio genetico. La differenza può essere determinata da fattori ambientali (lo vedremo meglio più avanti) o da una diversa predisposizione determinata da geni diversi. Resta il fatto che, in questo caso, il fenotipo si manifesta in modo non completamente dipendente dal solo patrimonio genetico. Per esempio, è pressoché impossibile stabilire con esattezza la distribuzione della pezzatura. Le piccole macchie non si trasmettono come fossero tante piccole mutazioni a sé. Soggetti con lo stesso patrimonio genetico possono presentare diversi disegni, perché in un individuo può prevalere il mantello solido e nell’altro quello pezzato, ferme restando le differenze determinate dall’azione dei geni modificatori.
Un altro esempio a testimonianza di questo tipo di eredità è la displasia dell’anca. Questa malattia, che si manifesta producendo una sublussazione coxofemorale, è teoricamente recessiva. Molti soggetti eterozigoti sono però sono riconoscibili all’esame radiografico e, a volte, manifestano i sintomi della malattia solo in età avanzata. Ve ne sono altri però che, fenotipicamente, sono perfettamente sani. Eppure trasmettono la malattia ai figli. Anche in questo caso vi sono fattori esterni e ambientali che possono essere determinanti. Difficile dire quanto dipenda dalla genetica e quanto da altri fattori.
L’epistasi
Con questo termine intendiamo l’effetto col quale geni non allelici possono mascherare la reciproca espressione fenotipica. Nell’esempio precedente, abbiamo visto che la feomelanina può assumere diverse tonalità, dal crema al fulvo carico. I geni responsabili di queste caratteristiche però non riescono a manifestare i loro effetti quando un gene del tutto estraneo (non allelico) interferisce bloccando l’espressione della feomelanina e trasformandola in eumelanina. E’ questo il caso del melanismo completo che determina il colore nero, marrone o grigio esteso uniformemente in tutte le aree pigmentate. Ogni cane nero, pur non manifestandolo, conserva nel proprio patrimonio genetico i geni responsabili della tonalità del colore fulvo ed è in grado a trasmetterli alla discendenza.
I geni letali
Sono geni che possono si esprimono solamente allo stato di eterozigosi. I soggetti omozigoti muoiono prima ancora della nascita. Un tipico gene letale, per esempio è quello che provoca l’assenza del pelo nelle razze di cani nudi (Chinese Crested Dog, Cane Nudo Peruviano, Xoloitzcuintle, ecc.). Accanto a soggetti nudi, troviamo regolarmente altri con mantello normale. I cani nudi, sono infatti portatori di mantello normale, mentre eventuali omozigoti per il gene “nudo”, morirebbero prima della nascita.
Nn x Nn =
25% NN: non vitali
50% Nn: nudi
25% nn: con mantello normale
Per questa ragione, in un programma razionale d’allevamento, in presenza di questo gene, si consiglia sempre di accoppiare puro per portatore, in modo da ottenere prole interamente vitale di cui un 50% di puri e un 50% di portatori;
Un altro esempio di gene letale è il fattore M «Merle». Questo gene, per la precisione, è subletale. La morte sembra essere più frequente in presenza di mantelli pezzati. Nei mantelli solidi, comunque è abbastanza frequente che gli omozigoti MM abbiano gravi problemi di salute (cecità, sordità, poca, costituzione debole).
Il mosaicismo
Capita talvolta di vedere cani a macchie nere su fondo biondo o altre disposizioni infrequenti dei colori. Questi mantelli sono probabilmente da attribuire a quel fenomeno che in genetica prende il nome di mosaicismo. Significa, che in una delle primissime divisioni dello zigote, vi è stata un’irregolarità che si è trasmessa solo alle cellule derivate da questa. In particolare, se l’irregolarità è avvenuta alla primissima divisione meiotica, avremo un cane che presenterà la metà destra di un colore e la metà sinistra di un colore diverso. La prima divisione è infatti quella che determina le due cellule dalle quali, nelle successive divisioni si formeranno rispettivamente tutte le cellule della metà destra e tutte quelle della metà sinistra del corpo.
I geni e l’ambiente
L’interazione e la dominanza non sono le uniche cause che determinano la variazione dell’espressione fenotipica di un gene. Possono intervenire anche altri fattori, dipendenti soprattutto dall’ambiente esterno. L’effetto dell’ambiente non è sempre uguale su tutti gli individui. Come ho già spiegato, parlando della penetranza incompleta, in presenza di determinate condizioni ambientali esterne o di una determinata alimentazione, può succedere che solo alcuni individui aventi lo stesso genotipo manifestino un certo fenotipo. L’effetto ambientale sul fenotipo è spesso irreversibile. Si pensi all’altezza. E’ provato che cani provenienti dallo stesso ceppo genetico, sottoposti a tipi diversi di alimentazione possono sviluppare altezze diverse, come diversa può essere la struttura scheletrica. Una volta adulti, un successivo cambio di alimentazione non porterà alcuna modificazione dell’altezza raggiunta. Tuttavia l’altezza determinata da questi fattori resta comunque un fattore non ereditario e si limitata a una sola generazione.
Denis Ferretti